Transducing mechanical cues
 |
Cells are embedded in mechanical cues, such as pulls, distortions, compressions and stretching. The cell's own shape is a "balancing act" of different forces: for example, the cell resists to external mechanical stress by adjusting the tension and organization of its cytoskeleton and adhesion sites. Tissue mechanics shapes organs during development and contributes to their homeostasis. Aberrant changes in tissue structure and mechanics, or in the cellular response to mechanical stresses, are a general feature of most human diseases (including cancer, fibrosis and vascular diseases). And accumulating evidence indicates that altered cell and tissue distortions actually anticipate overt disease and play a causal role in disease etiology and progression.
We pioneered the notion that mechanical signals and cell shape are converted into biochemical responses by two related transcription factors, YAP and TAZ (Dupont et al., Nature 2011; Halder et al., Nat. Rev. MCB 2012). We found that changes in cell shape caused by i) attachment to stiff vs. soft extracellular matrix (ECM) substrates; ii) confined adhesion to big vs. small "islands" of a microprinted substrate; iii) changes in cytoskeletal organization, all cause YAP and TAZ to enter the nucleus and activate their targets (in stiff environments, spread-out cells, high cytoskeletal tension), or exit the nucleus (in cells that are soft, small, and with a relaxed actin cytoskeleton). As such, these studies add an additional dimension to cell signaling, whereby individual cells can be informed about properties of the tissue in which they are embedded, such as its size and three-dimensional organization (Dupont et al., Nature 2011; Halder et al., Nat. Rev. MCB 2012).
Questions we are currently addressing include: what are the molecular players in the mechanotransduction pathways linking integrins, cell-cell adhesion complexes and the actin cytoskeleton to TAZ and YAP activity? Is the geometry of ECM and tissue mechanics a source of positional information through YAP and TAZ? Can one normalize aberrant cell behavior in disease setting by "normalizing" cell mechanics?
Please click here for general advice and technical tips on how to monitor YAP/TAZ regulation and activity in cell culture by 8xGTIIC lux.
For protocols on testing the response of cells to mechanical stimuli, click here
We pioneered the notion that mechanical signals and cell shape are converted into biochemical responses by two related transcription factors, YAP and TAZ (Dupont et al., Nature 2011; Halder et al., Nat. Rev. MCB 2012). We found that changes in cell shape caused by i) attachment to stiff vs. soft extracellular matrix (ECM) substrates; ii) confined adhesion to big vs. small "islands" of a microprinted substrate; iii) changes in cytoskeletal organization, all cause YAP and TAZ to enter the nucleus and activate their targets (in stiff environments, spread-out cells, high cytoskeletal tension), or exit the nucleus (in cells that are soft, small, and with a relaxed actin cytoskeleton). As such, these studies add an additional dimension to cell signaling, whereby individual cells can be informed about properties of the tissue in which they are embedded, such as its size and three-dimensional organization (Dupont et al., Nature 2011; Halder et al., Nat. Rev. MCB 2012).
Questions we are currently addressing include: what are the molecular players in the mechanotransduction pathways linking integrins, cell-cell adhesion complexes and the actin cytoskeleton to TAZ and YAP activity? Is the geometry of ECM and tissue mechanics a source of positional information through YAP and TAZ? Can one normalize aberrant cell behavior in disease setting by "normalizing" cell mechanics?
Please click here for general advice and technical tips on how to monitor YAP/TAZ regulation and activity in cell culture by 8xGTIIC lux.
For protocols on testing the response of cells to mechanical stimuli, click here
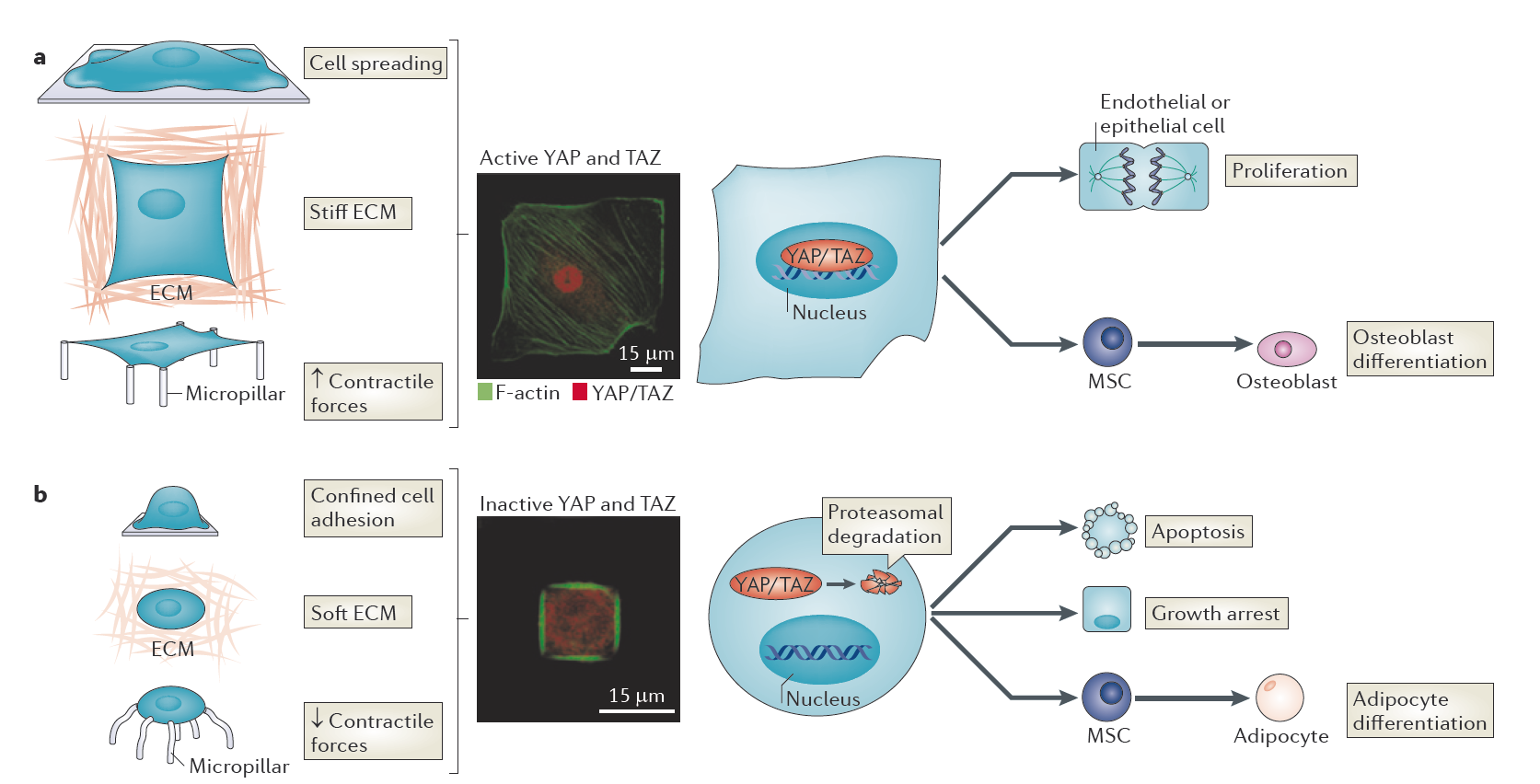 YAP and TAZ as sensors and mediators of mechanical inputs from the extracellular matrix. a | YAP and TAZ are localized in the nucleus and active under experimental mechanical conditions that favour the development of high intracellular resisting forces, such as an unlimited adhesive area and the subsequent adoption of a spread cell shape, a stiff extracellular matrix (ECM) or stretching between stiff micropillars (left). Green: Phalloidin-staining for filamentous actin (F-actin); Red: YAP and TAZ. b | In mechanical conditions in which cells develop low contractile forces (for example, when they are grown on small adhesive areas favouring a small cell size, on soft ECM or on top of bendable micropillars), YAP and TAZ are inactivated and relocalize to the cytoplasm. Reduced YAP and TAZ activity shifts the cell responses towards apoptosis and growth arrest or diverts differentiation towards cell fates that would be specified on a soft matrix, for example, adipocytes. (From Halder, Dupont and Piccolo, 2012). |
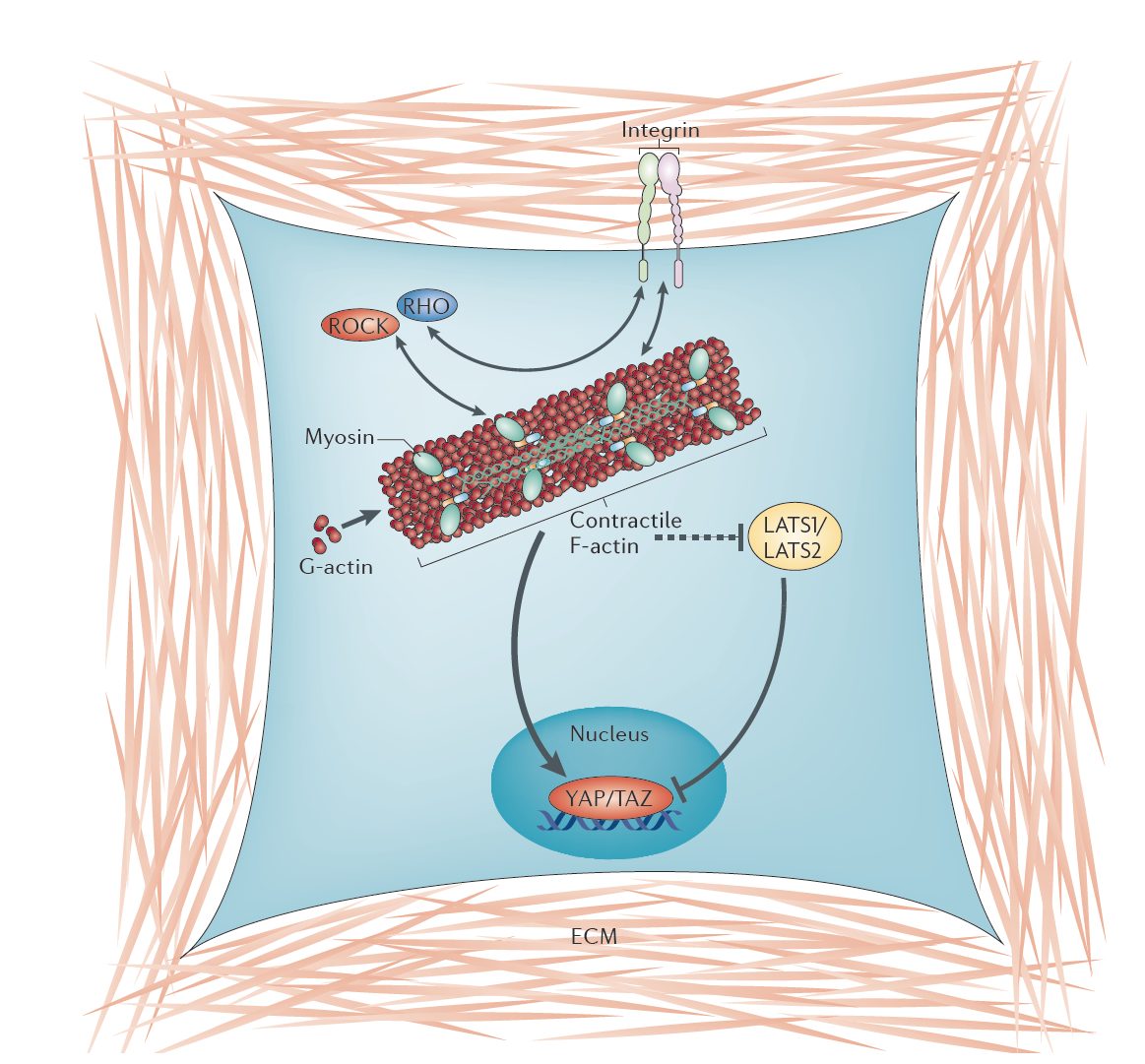 A model for the mechanical regulation of YAP and TAZ. When cells are
allowed to spread freely on the extracellular matrix (ECM), or when
cells are grown on a stiff ECM, extracellular forces promote cell-ECM
adhesions via integrins and the development of intracellular
contractile filamentous actin (F-actin) structures containing myosin
molecules. This is regulated by bidirectional signalling between RHO,
ROCK (RHO-associated kinase), integrins and myosin activity.
Contractile F-actin structures in turn sustain YAP and TAZ nuclear
localization and activity through unidentified molecular effectors.
In addition, F-actin also opposes YAP and TAZ phosphorylation through
inhibition of the kinases LATS1 (Large tumour suppressor homologue 1)
and LATS2. G-actin, globular actin. (From Halder, Dupont, Piccolo 2012). |
|
click the images to enlarge | ||